

Todo el mundo sabe que el coste de una obra de teatro además del trabajo de los actores incluye a mucho personal entre bambalinas. El coste termodinámico de una ruta metabólica también incluye un coste entre bambalinas, el coste proteómico de la síntesis de sus enzimas. Nos lo recuerda un nuevo artículo en PNAS que estudia en procariotas la ruta metabólica de la glucólisis, la síntesis de energía (moléculas de ATP) oxidando glucosa. La ruta metabólica estándar para la glucólisis es la de Embden-Meyerhoff-Parnass (EMP), que contiene 10 reacciones enzimáticas y cuyo producto son dos moléculas de ATP. Sin embargo, hay otras rutas para la glucólisis en procariotas, siendo la más común la ruta de Entner-Doudoroff (ED), que produce una sola molécula de ATP, pero sólo tiene 6 reacciones enzimáticas. Avi Flamholz (Instituto Weizmann de Ciencia, Rehovot, Israel) y sus colegas afirman que la prevalencia de la ruta ED en procariotas es debido a que requiere menos enzimas (6 en lugar de 10); este resultado lo han obtenido tras un análisis termodinámico de la cinética química de estas rutas metabólicas que incluye el coste termodinámico de la producción de las enzimas (normalmente despreciado en este tipo de análisis). El nuevo estudio nos recuerda que las bambalinas también existen en el metabolismo celular. Nos lo cuentan Arion I. Stettner, Daniel Segrè, «The cost of efficiency in energy metabolism,» PNAS, AOP May 31, 2013, quienes se hacen eco del artículo técnico de Avi Flamholz et al., «Glycolytic strategy as a tradeoff between energy yield and protein cost,»PNAS, AOP Apr 29, 2013.
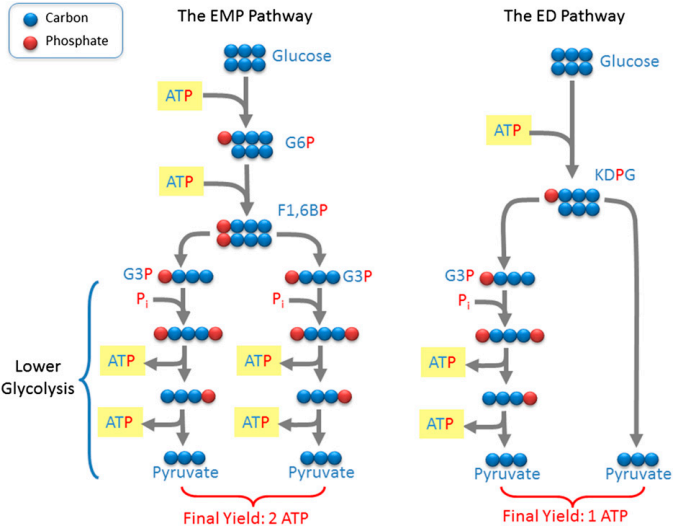
La estequiometría química de las rutas metabólicas de Embden-Meyerhof-Parnas (EMP) y Entner-Doudoroff (ED) parece indicar que la ruta EMP es más óptima que la ruta ED ya que produce dos veces más ATP para la misma cantidad de glucosa. En la primera etapa de la ruta EMP, la glucosa es fosforilada dos veces, consumiendo dos ATP, mientras que en ruta ED sólo es fosforilada una vez. Se cree por ello que la ruta ED es filogenéticamente más antigua que la ruta EMP (de hecho, es la dominante entre los eucariotas). Además, se cree que la ruta EMP es un refinamiento evolutivo de la ruta ED para optimizar su rendimiento (o eficiencia). Sin embargo, esta optimización tiene un costo enzimático mucho mayor.
Flamholz et al. miden el coste enzimático en cada ruta metabólica utilizando la relación de Haldane aplicada a la cinética enzimática de Michaelis-Menten; esta relación muestra de forma explícita la cantidad de enzima necesaria para la reacción química. Han aplicado un algoritmo de optimización con restricciones (cuyo código está diponible en la web y aparece en la información suplementaria de acceso gratuito) para demostrar que la ruta de EMP tiene un coste en proteínas 3,5 veces mayor que la ruta ED. Este mayor coste enzimático conlleva un alto consumo de proteoma (algo ya reconocido por autores anteriores en Escherichia coli y Saccharomyces cerevisiae). Como la ruta EMP tiene una alta producción de ATP a costa de una costosa maquinaria enzimática, nos podemos preguntar ¿por qué los procariotas prefieren la ruta ED y los eucariotas la EMP? Como siempre, la dinámica evolutiva conlleva ciertos compromisos y el coste enzimático es más relevante en seres vivos más simples (como los procariotas), en los que el efecto de la epistasis es menor.
En mi opinión, el nuevo trabajo de Flamholz et al. nos recuerda que no siempre lo que está más a la vista es lo más relevante y que los enfoques holísticos en biología de sistemas nos van a ofrecer gran número de sorpresas a la hora de entender la complejidad del metabolismo de los seres vivos.
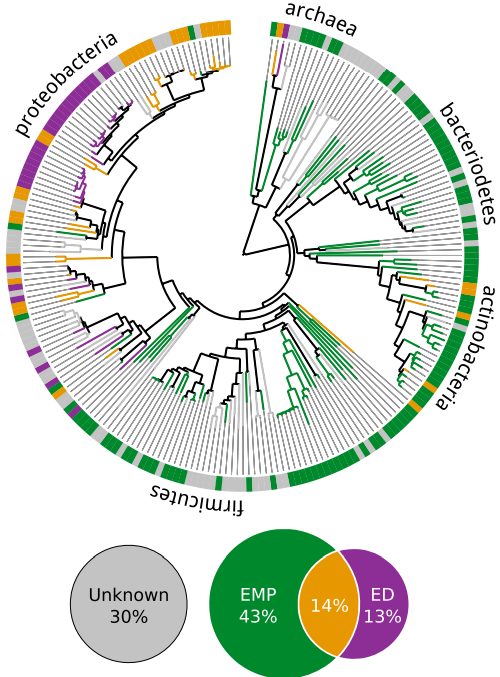
Como se ve en la figura 6 del artículo:
http://www.pnas.org/content/early/2013/04/26/1215283110.full.pdf+html
los anaerobios usan prácticamente siempre (97%) la ruta EMP, que rinde 2 ATP, pero como los dos piruvatos producidos no pasan al ciclo de Krebs, ya que este ciclo sólo funciona acoplado a la cadena respiratoria, es decir que se necesita oxígeno para que sea eficaz, esos 2 ATP son el mejor rendimiento que pueden obtener; mientras que en los aerobios casi la mitad (45%) usan la ruta ED, ya que aunque solo produce 1 ATP, los piruvatos pueden ser convertidos por descarboxilación en acetilCoA, que pasa al ciclo de Krebs y cadena respiratoria, donde producirá 12 ATP. Por tanto para los anaerobios no tiene sentido usar la ruta ED, mientras que para los aerobios sí.
Yo creo que los eucariotas utilizan la ruta EMP porque en ella esta presente la enzima fosfofructoquinasa 1 (PFK 1) que le «añade» un nuevo grupo fosfato (C1 exactamente). Esta enzima cataliza la reacción que es controlador de flujo de toda la glicólisis ya que es muy exergonica. Gracias a esta enzima se puede controlar la glicólisis de una forma mucho más compleja (cosa que en eucariotas viene mas bien que en bacterias y arqueas me parece a mi). Por ejemplo, la fructosa 2,6-bisfosfato es la principal reguladora (regulación alosterica) de esta enzima la cual se puede desfosforilar a glucosa 6 fosfato o no según convenga ya que los enzimas que la desfosforilan (glucosa 6 fosfato) y los que las fosforilan ( glucosa 2,6-bisfosfato) están regulados a la vez por los productos de las hormonas metabólicas como la insulina o el glucagon. Esto permite tener una regulación del metabolismo a nivel global y no solo de célula por célula (cosa que viene bien en organismos pluricelulares complejos). Es una opinión cualquiera xd.
Me parece que el balance de la vía ED debe verse diferente. 1NADPH equivale a 1 NADH + 1 ATP. Haciendo esta consideración ambas rutas decantan 2 piruvatos, 2 NADH y 2 ATP. Desconozco si los estrictos anaerobios requieren NADPH para las reducciones anabólicas. Si fuese así, aquí sería el momento de pasárseles la factura y tener que invertir un ATP para convertir NADH en NADPH.