


La ley de Kleiber (1932) afirma que el metabolismo basal (B) en mamíferos sigue una ley de potencia con su masa (M) de exponente 3/4, es decir, B = c M3/4. En muchos organismos, como aves e insectos, la ley falla y se requiere un exponente entre 2/3 y 1. Fernando J. Ballesteros (Univ. València, España) y varios colegas generalizan en Scientific Reports esta ley mediante una suma de dos leyes de potencia, B = a M + b M2/3. Su idea es interpolar la disipación calórica pasiva, el término alométrico B = b M2/3, y el gasto energético mínimo por célula, el término isométrico B = a M. La ventaja de la nueva ley generalizada es que da cuenta de la curvatura observada en ciertos estudios empíricos (LCMF, 31 Mar 2010).
El artículo es Fernando J. Ballesteros, Vicent J. Martinez, …, Andrés Moya, «On the thermodynamic origin of metabolic scaling,» Scientific Reports 8: 1448 (23 Jan 2018), doi: 10.1038/s41598-018-19853-6. He de confesar que el primer autor, Fernando J. Ballesteros, me envió la nota de prensa de su universidad por e-mail hace unos días; pensó que me podría interesar tras la lectura de «La relación entre la tasa metabólica y la masa corporal no es una ley de potencias en mamíferos», LCMF, 31 Mar 2010, donde me hacía eco de Tom Kolokotrones, Van Savage, …, Walter Fontana, «Curvature in metabolic scaling,» Nature 464: 753–756 (01 Apr 2010), doi: 10.1038/nature08920. Puedes leer la nota de prensa en «Investigadores de la Universitat descubren por qué el metabolismo basal de los seres vivos varía con su masa», UV, 24 Ene 2018; también «Resuelto un antiguo enigma biológico. Aclarado por qué el metabolismo de los animales depende de su masa», Agencia SINC, 26 Ene 2018.
También he de confesar que lo que más me sorprendió de la nota de prensa es la divertida imagen de una persona con paraguas rosa en la figura que la acompaña (y que abre esta entrada). Sin lugar a dudas parece que está haciendo el ganso, frente a un ganso, bajo la atenta mirada de un incrédulo perro. No sé qué pensarás, pero estos toques de humor en las notas de prensa siempre son de agradecer; por fortuna no aparece en el artículo científico, donde me parecería una nota de mal gusto.
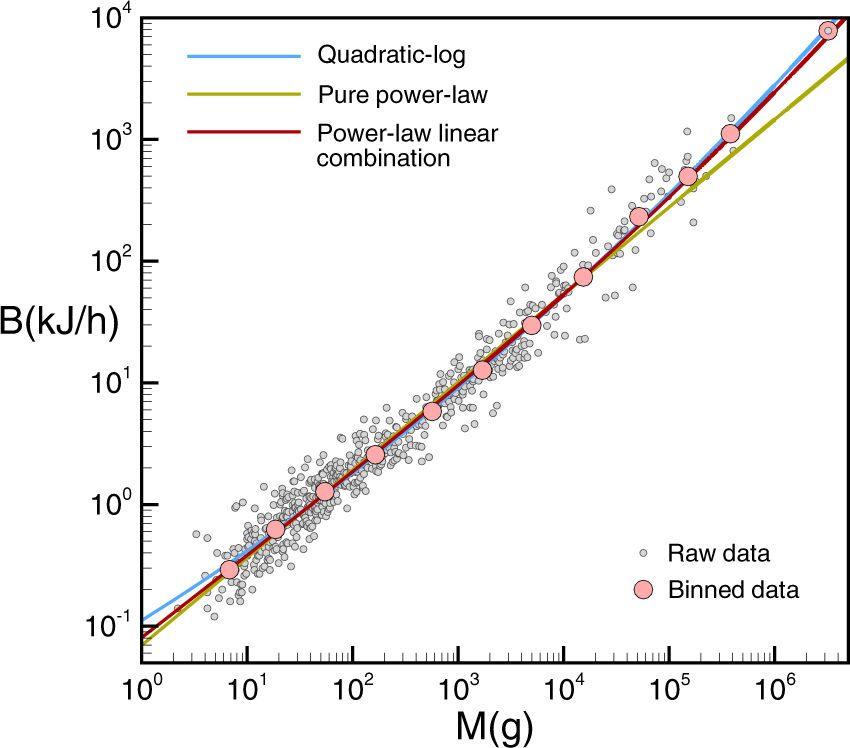
El metabolismo basal B (kJ/h, kilojulios por hora) es la energía mínima que consume un organismo para mantenerse vivo; la ley de Kleiber lo relaciona con la masa M (g, gramos). Esta figura muestra los datos estimados por McNab para 637 mamíferos (B. K. McNab, «An analysis of the factors that influence the level and scaling of mammalian BMR,» Comparative Biochemistry and Physiology A—Molecular & Integrative Physiology 151: 5-28 (2008), doi: 10.1016/j.cbpa.2008.05.008; por cierto, todos estos datos para mamíferos los tienes en este fichero Excel). En esta figura la ley de Kleiber es la curva verde, la ley cuadrática propuesta por Kolokotrones et al. en 2010 es la curva azul y la nueva ley de Ballesteros et al. es la curva roja. Como puedes observar el ajuste entre estas dos últimas es bastante bueno, luego la nueva ley incorpora el efecto de la curvatura publicado por Kolokotrones et al. en Nature.

Parte de esta energía se usa para el funcionamiento del organismo y parte se disipa en forma de calor. Si toda esta energía se transformara en calor, el organismo sería una estufa con B = b M2/3. Si las células consumieran toda esta energía con una eficiencia máxima, el consumo sería proporcional a su número, es decir, su masa, B = a M. En la práctica los organismos interpolan entre ambos extremos, B = f k M + (1−f) k‘ M2/3, donde k y k‘ son los coeficientes de Meeh. Estos coeficientes varían según los organismos y el artículo estudia cómo varía el cociente b/a ∝ (1 − f)/f entre mamíferos desérticos y polares, entre mamíferos, aves e insectos, y entre animales y plantas.

En mi opinión, lo más interesante del nuevo artículo es que propone un modelo termodinámico para estimar los coeficientes de Meeh. En concreto, k‘ = d (hR+hC) ΔT ρ−2/3, donde d es un parámetro adimensional que depende de la geometría del cuerpo, hR y hC son los coeficientes de transferencia térmica por radiación y convección, resp., ΔT = Ts − Te es la diferencia entre la temperatura del cuerpo Ts y la del entorno Te, y ρ es la densidad del cuerpo; un valor típico para mamíferos es k’ ≈ 0,1, que como b = k’ (1 − f) ≈ 0,079, resulta f ≈ 0,21 (luego k = a/f ≈ 0,0076 kJ/hg). Este valor es razonable ya que se estima que solo el 42% de la energía almacenada en una molécula de glucosa se captura en forma de ATP (el 58% se disipa en forma de calor), el coeficiente f ≤ 0,42, y que el 50% del ATP se consume en reacciones biológicas, luego se espera f ≈ 0,21.
Por supuesto, el nuevo artículo no se libra de las críticas típicas a todas las estimaciones de datos empíricos mediante leyes de potencia. Como suele ser habitual, Ballesteros et al. usa un método de estimación sesgado (regresión lineal mediante mínimos cuadrados aplicada al logaritmo de los datos). Este método produce resultados poco robustos, que cambian mucho bajo pequeñas modificaciones de los datos (en especial bajo datos espurios). Yo no soy experto en estadística, pero hubiera un estimador de máxima verosimilitud, por ejemplo, siguiendo a Aaron Clauset, Cosma Rohilla Shalizi, M. E. J. Newman, «Power-law distributions in empirical data,» SIAM Review 51: 661-703 (2009), doi: 10.1137/070710111, arXiv:0706.1062 [physics.data-an]), artículo que recomendé en «Todo lo que siempre quisiste saber sobre leyes de potencia», LCMF, 21 Jun 2009. O quizás hubiera aplicado a los datos B/M = a + b M−1/3 el método propuesto en Rudolf Hanel, Bernat Corominas-Murtra, …, Stefan Thurner, «Fitting power-laws in empirical data with estimators that work for all exponents,» PLoS ONE, 28 Feb 2017, doi: 10.1371/journal.pone.0170920.
Por otro lado, sustituir una ley de potencias por una suma de dos leyes de potencia introduce grandes dificultades en la interpretación estadística de los datos (la suma de leyes de Pareto no es una ley de Pareto y no debe ser interpretada en los mismos términos; ver, por ejemplo, Colin M. Ramsay, «The Distribution of Sums of Certain I.I.D. Pareto Variates,» Communications in Statistics – Theory and Methods 35: 395-405 (2006), doi: 10.1080/03610920500476325). Si bien el nuevo artículo de Ballesteros et al. omite un análisis detallado de la interpretación biológica de la nueva ley generalizada propuesta, en mi opinión, la mayoría de los fisiólogos (a quienes va dirigido el tragajo) tendrán grandes dificultades a la hora de interpretar la nueva ley para las colas de la distribución (donde su uso es más sugerente); nunca deben olvidar que aplicar la interpretación estadística convencional de la ley de Kleiber les llevará a sinsentidos desde el punto de vista estadístico.
En resumen, un artículo interesante pero que creo que generará críticas entre fisiólogos y estadísticos (lo que les dará citas a los autores). Sin lugar a dudas, las leyes de potencia ubicuas, pero su uso en ciencia está repleto de riesgos. Quienes se atrevan a extraer conclusiones a partir de la nueva ley generalizada de Kleiber tendrán que ser muy cautos en este sentido.
A todas luces, el tipo del paraguas representa mucho mejor a los humanos que el simple bosquejo del hombre o mujer, desnudos, mirando al frente…vamos, no hay color.
Representa estupendamente en qué gasta calorías el ser humano 🙂
Creo recordar que ya vi la figura del paraguas rosa en una charla de Geoffrey West en Santa Fe.
Pues me la debió de copiar él a mí 😉 porque esa la elaboré yo desde cero para este libro: «Caos y Fractales» (Guadalmazan, 2017) aunque allí con la ley de Kleiber canónica. Para la nota de prensa cambié la power law por la curva que da nuestro modelo.
«Como suele ser habitual, Ballesteros et al. usa un método de estimación sesgado (regresión lineal mediante mínimos cuadrados aplicada al logaritmo de los datos).»
No es del todo correcto: la regresión es no lineal (en log-log el modelo del fit no es lineal, pues una combinación de leyes de potencia no es una ley de potencia). El método es adecuado pues observamos que en espacio logarítmico, observamos que la dispersión de los datos era Gaussiana.
Gracias, Lucas, por la aclaración (como coautor no esperaba menos). Por cierto, si construyes una población de datos eliminando, pongamos, el 10% de tus datos, y repites tu regresión «no lineal» observarás una fuerte correlación líneal entre los parámetros estimados, espacio (a,b), como es de esperar para una regresión «lineal», aunque ejecutada con métodos no lineales.
Muy interesante. Viendo las gráficas del artículo y la dispersión que tienen los datos, aparte de que el ajuste se pueda hacer algo mejor o peor, creo que lo más que se puede decir es que es compatible con los datos. Como muchos otros modelos, supongo. La cuestión sería ver si este modelo tiene más sentido que sus competidores, si queda mejor parado con la navaja de Ockham. O sea, si tienes dos modelos A y B, ambos compatibles con los datos, y A ajusta algo mejor que B pero a la vez es muchísimo más complejo, yo preferiría el B.
Excelente artículo, como siempre. Es impresionante ver como se funden la mejor divulgación científica con una revisión por pares.
Hay una ecuacion para determinar el metabolismo basal del cuerpo humano desde el punto de vista termodinámico. Por si a,alguien le interesa.